CANC…RISATION ET EXPRESSION DES G»NES
Beljanski poursuit ses expťriences, ťclairť par la lumiŤre de son interprťtation nouvelle.
Comme d'habitude, l'ťquipe aborde parallŤlement plusieurs voies de recherche, que
nous
devrons dťcrire dans un ordre arbitrairement choisi. Avant de consacrer l'attention
qu'elle
mťrite ŗ la mise au point d'anticancťreux spťcifiques, nous voudrions relater un
certain nombre
de travaux qui concernent divers aspects de la dťstabilisation de 1'ADN.
I1 faut d'abord parler de la solution apportťe aux ťnigmes du test d'Ames: pourquoi
des
cancťrogŤnes de nature chimique trŤs diffťrente provoquent-ils tous, chez Salmonella
typhimurium, la mÍme mutation de la souche his- en his+ et pourquoi 20 % des substances
dont
l'action cancťrogŤne est dťmontrťe chez l' animal se montrent-elles incapables de
causer cette
mutation ?
Beljanski postule que 1'ADN des bactťries his- est. sous l'action des cancťrogŤnes,
beaucoup
plus fortement dťstabilisť que celui des bactťries sauvages his+ parce qu'il est. ŗ
l'origine, dťjŗ
dťstabilisť. I1 devrait donc se comporter dans l'oncotest exactement comme les ADN
cancťreux de mammifŤres, tandis que 1'ADN de la souche sauvage devrait rťagir comme
tout
ADN normal. C'est effectivement ce que les expťriences de l'ťquipe confirment.
En outre, quand 1'ADN isolť ŗ partir de la bactťrie mutante his- est utilisť comme
matrice dans
l'oncotest, il donne, cette fois, des rťponses positives sous l'effet des cancťrogŤnes
qui ne sont
pas dťtectťs, dans le test d' Ames, au niveau de la bactťrie entiŤre. I1 est
vraisemblable que ces
substances, parmi lesquelles de puissants cancťrogŤnes comme l'ťthionine, ne sont pas
dťtectables parce qu'elles ne peuvent tout simplement pas franchir la membrane
bactťrienne.
La dťstabilisation de 1'ADN de S. typhimurium his~ le rend vulnťrable. On peut
supposer qu'il
existe, au niveau d'un ou de plusieurs gŤnes concernťs par la synthŤse de l'histidine,
des sites
privilťgiťs oý les liaisons hydrogŤne sont particuliŤrement exposťes ŗ se rompre
sous l'action
des cancťrogŤnes, ce qui rend possible l'activation du gŤne correspondant.
Sans doute peut-on rapprocher ce phťnomŤne du processus qui rend les tissus
vťgťtaux
anergiťs indťpendants d'un apport extťrieur d'hormone de croissance. La nature des
produits
anormaux synthťtisťs par les ADN cancťreux ou prťcancťreux sous 1' action d' agents
dťstabilisants, c'est-ŗ-dire l'activation de certains gŤnes plutŰt que d'autres, est
probablement
dťterminťe, essentiellement, par le degrť de fragilitť des liaisons hydrogŤne au
niveau des
diffťrents sites le long du gťnome.
I1 existe ainsi, dans les ADN cancťreux, un certain nombre de sites plus fragiles sur
lesquels
les molťcules dťstabilisantes peuvent se fixer successivement et indiffťremment: en
effet, dans
la plupart des cas, elles ne manifestent pas d'affinitť spťcifique pour l'un plutŰt
qu'un autre.
C'est ce que permettent de dťduire les expťriences de l'ťquipe qui dťmontrent
l'additivitť des
effets des substances dťstabilisantes. Les effets de composťs diffťrents
s'additionnent, ceux des
diverses concentrations ťgalement.
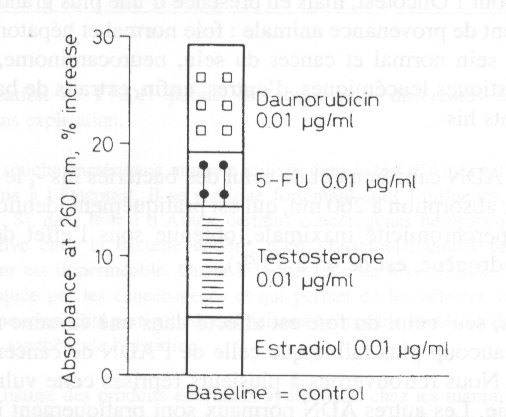
Dans l'une des ťpreuves destinťes ŗ dťmontrer cette additivitť, un ADN isolť ŗ
partir de cancer
du sein a ťtť incubť en prťsence de concentrations croissantes de diffťrents agents
dťstabilisants ajoutťs successivement au milieu d'incubation: d'abord des hormones
stťroÔdes,
l'estradiol, dont cet ADN est la cible physiologique, puis la testostťrone; ensuite, un
solvant
trŤs employť en laboratoire, le DMSO (dimťthylsulfoxyde); enfin, un antimitotique, la
daunorubicine. Les effets de ces produits s'additionnent, comme l'indiquent ŗ la fois la
mesure
de l'hyperchromicitť et l'ťvaluation de la quantitť de nouvel ADN synthťtisť. Chaque
substance contribue ŗ des degrťs diffťrents ŗ la sťparation locale des ch‚Ónes de
1'ADN et,
l'ťquipe l'a dťmontrť, il s'agit bien d'un ťcartement des brins, non de cassures.
< IMG_P47.JPG >
Sťparation des chaÓnes d'ADN du neurocarcinome. Effets de: estradiol,
testostťrone, 5-FU et daunorubicine. Absorbance ŗ 260 nm (rťf.88).
Les effets de ces produits s'ajoutent, comme en tťmoigne
l*ugmentation de l'hyperchromicitť (absorbance des Ud.
Un des composťs utilisťs dans cette expťrience, le DMSO, prťsente un intťrÍt
particulier.
Comme l'huile de croton (frťquemment employťe comme irritant de la peau chez l' animal
de
laboratoire, notamment pour favoriser l' apparition de tumeurs cutanťes), le DMSO induit
l'ouverture des ch‚Ónes d'ADN; mais ces deux substances ne font pas de distinction entre
1'ADN cancťreux et 1'ADN normal et se fixent ŗ l'un comme ŗ l'autre. Or de nombreuses
expťriences ont prouvť que le DMSO et l'huile de croton, ainsi que, du reste,
l'actinomycine
D, qui se comporte en cancťrogŤne dans l'Oncotest, ont la capacitť, ŗ certaines
concentrations,
de provoquer la diffťrenciation de divers types de cellules. Ce fait, constatť sans
Ítre interprťtť,
trouve dťsormais son explication: en ouvrant localement la chaÓne de 1'ADN, les agents
dťstabilisants permettent aux gŤnes de s'exprimer.
Une autre molťcule, le TPA (12-tťtradťcanoylphorbol-13-acťtate), exerce des effets
multiples
qui n' avaient pas non plus reÁu d' explication. TrŤs utilisť en biochimie, cet ester
isolť ŗ partir
de l'huile de phorbol se comporte gťnťralement comme un promoteur des tumeurs,
c'est-ŗ-dire
qu'il en accťlŤre la prolifťration. Nťanmoins, comme les autres promoteurs de tumeurs,
il peut
en outre, ŗ forte concentration, Ítre aussi inducteur de cancers.
Le TPA manifeste bien d' autres activitťs. I1 stimule la prolifťration des cellules
embryonnaires"'. I1 peut, comme le DMSO, induire la diffťrenciation des cellules
leucťmiques,
qui, d'habitude, demeurent dans un ťtat peu diffťrenciť. I1 peut aussi activer, dans
les
hybridomes'2', des gŤnes silencieux qui vont alors donner naissance aux ARN ribosomiques
de
(1) Parus en 1982, les travaux qui ont dťcrit la stimulation de la croissance des
cellules embryonnaires par le TPA
n'ťtaient pas encore connus au moment oý l'ťquipe menait ses expťriences; ils ne font
que confirmer le bien-
fondť de la nouvelle thťorie.
(2) Rappelons que les hybridones sont des cellules fommťes artificiellement, au
laboratoire, par la fusion d'un Iymphocyte
(qui a une durťe de vie lim itťe) et d' une cellule d'origine cancťreuse (qui se multi
pl ie indťfiniment en culture).
ces cellules-chimŤres. Enfin, le TPA peut modifier le phťnotype - le rťsultat
visible du travail
des gŤnes - de certaines espŤces cellulaires.
Le TPA et, pour servir de tťmoin, un autre dťrivť de l' ester de phorbol, l'
alpha-phorbol, qui est
dťpourvu d' action sur la croissance cellulaire et la tumorogenŤse, ont ťtť incubťs
in vitro selon
la mťthode utilisťe pour l'Oncotest, mais en prťsence d'une plus grande variťtť
d'ADN; parrni
ceux-ci, les uns ťtaient de provenance animale: foie normal et hťpatome greffť de rat,
d'autres
d'origine humaine: sein normal et cancer du sein, neurocarcinome, leucocytes normaux et
leucocytes myťloblastiques leucťmiques, d'autres, enfin, extraits de bactťries: S.
typhEmurium
ainsi que trois mutants his-.
Mis en prťsence des ADN cancťreux et de celui des bactťries his -, le TPA provoque
une forte
augmentation de leur absorption ŗ 260 nm, qui est pratiquement identique pour tous: 22 ŗ
25 %
(rappelons que l'hyperchromicitť maximale, obtenue sous l'effet de la potasse, qui rompt
toutes les liaisons hydrogŤne, est de 45 ŗ 55 %).
Parmi les ADN sains, seul celui du foie est affectť dans une certaine mesure:
l'ouverture des
ch‚Ónes, bien que beaucoup plus faible que celle de 1'ADN de cancer du foie, est
nťanmoins
aisťment dťtectable. Nous retrouverons ŗ plusieurs reprises cette vulnťrabilitť
particuliŤre de
1'ADN normal de foie. Les autres ADN normaux sont pratiquement insensibles ŗ l'action du
TPA. Les rťsultats des mesures de la quantitť d'ADN synthťtisť vont exactement dans le
mÍme
sens. L' alpha-phorbol, dťnuť d' activitť sur la multiplication et la diffťrenciation
cellulaires, n' a
d'effet sur aucun des ADN testťs.
Le cas du TPA illustre bien la maniŤre dont la nouvelle thťorie permet de rattacher
des
observations jusque-lŗ sans lien et de les expliquer.
R…SUM…
La dťstabilisation de l'ADN permet de comprendre diffťrentes observations
demeurťes sans explication.
L'ADN de la souche bactťrienne mutante utilisťe dans le test d' Ames est
dťstabilisť.
Isolť et soumis ŗ l'Oncotest, il se rťplique fortement sous l'influence de tous les
cancťrogŤnes. Si , dans le test d' Ames, certains cancťrogŤnes ne provoquent pas de
rťponse positive chez la bactťrie entiŤre, c'est vraisemblablement parce que sa
membrane leur est impermťable. Quant ŗ la mutation de his - en his+, toujours la
mÍme, provoquťe par les cancťrogŤnes et qui permet de les dťtecter, elle est sans
doute due ŗ une sensibilitť particuliŤre des liaisons hydrogŤne au niveau des gŤnes
qui
gouvernent la synthŤse de l'histidine.
De mÍme, la nature des produits anormaux synthťtisťs, chez les mammifŤres et les
plantes, par les ADN cancťreux ou prťcancťreux sous l'action d'agents dťstabilisants
est dťterminťe essentiellement par le degrť de fragilitť des liaisons hydrogŤne au
niveau des diffťrents gŤnes.
Les cancťrogŤnes se fixent successivement et indiffťremment ŗ ces sites fragiles.
Des
expťriences de l'ťquipe dťmontrent que les substances dťstabilisantes, ainsi que leurs
diverses concentrations, ont des effets additifs.
Les multiples actions au niveau cellulaire de deux produits largement utilisťs en
biochimie, le DMSO et le TPA, notamment l'induction de la diffťrenciation,
s'expliquent par leur capacitť de dťstabiliser l'ADN.